Relaxin for the research of RXFPs bioactivity
Abstract
Tevz G, Mcgrath S, Demeter R, et al. Identification of a novel fusion transcript between human relaxin-1 (RLN1) and human relaxin-2 (RLN2) in prostate cancer. Mol Cell Endocrinol. 2016;420:159-68.
Follicle-stimulating hormone (FSH) stimulates the proliferation of immature Sertoli cells through the activation of PI3K/AKT/mTORC1 and MEK/ERK1/2 pathways. Mature Sertoli cells stop proliferating and respond to FSH by stimulating cAMP production. To gain insight into possible mechanisms involved in this switch as well as the impact of paracrine factors that stimulate cell proliferation, we analyzed the effects of FSH and relaxin on intracellular signaling pathways involved with proliferation and differentiation in Sertoli cells from 15-day-old rats, which are close to the transition between the two stages. FSH stimulated 3H-thymidine incorporation and cyclin D1 expression, changes associated with proliferation. In contrast, FSH inhibited AKT and ERK1/2 phosphorylation, activated cAMP production and induced changes in several cell cycle genes that were compatible with differentiation. Relaxin also stimulated 3H-thymidine incorporation but increased phosphorylation of ERK1/2 and AKT. When both hormones were added simultaneously, relaxin attenuated FSH-mediated inhibition of ERK1/2 and AKT phosphorylation and FSH-mediated activation of cAMP production. FSH but not relaxin increased CREB phosphorylation, and relaxin but not FSH shifted NF-κB expression from the cytoplasm to the nucleus. Relaxin did not inhibit the effects of FSH on inhibin α and Bcl2 expression. We propose that at this time of Sertoli cell development, FSH starts to direct cells to differentiation through activation of cAMP/CREB and inhibition of ERK1/2 and AKT pathways. Relaxin counteracts FSH signaling through the inhibition of cAMP and activation of ERK1/2, AKT and NF-κB, but does not block the differentiation process triggered by FSH.
This publication used human relaxin-2 peptide for intracellular signaling pathways study.
Nascimento AR, Macheroni C, Lucas TF, Porto CS, Lazari MF. Reproduction. 2016;152(6):613-628.
this Publication used the human relaxin-2 RIA Kit (RK-035-62) from Phoenix Pharmaceuticals for relaxin-2 serum sample level measurement.
Cernaro V, Lacquaniti A, Giorgianni G, Bolignano D, Buemi M. Modifications in relaxin’s serum levels during acetate-free biofiltration (AFB): only a new biomarker?. Ital J Anat Embryol. 2013;118(1 Suppl):98-9.
This Publication used a Relaxin-2 peptide (035-62) from Phoenix Pharmaceuticals.
Carvalho LN, Cristovam PC, Passos CS, Boim MA. Mesangial cells cultured from pregnant rats display reduced reactivity to angiotensin II: the role of relaxin, nitric oxide and AT2 receptor. Cell Physiol Biochem. 2012;30(6):1456-64.
Human relaxin-2 (hereafter simply defined as “relaxin”) is a 6-kDa peptidic hormone best known for the physiological role played during pregnancy in the growth and differentiation of the reproductive tract and in the renal and systemic hemodynamic changes. This factor can also be involved in the pathophysiology of arterial hypertension and heart failure, in the molecular pathways of fibrosis and cancer, and in angiogenesis and bone remodeling. It belongs to the relaxin peptide family, whose members comprehensively exert numerous effects through interaction with different types of receptors, classified as relaxin family peptide (RXFP) receptors (RXFP1, RXFP2, RXFP3, RXFP4). Research looks toward the in-depth examination and complete understanding of relaxin in its various pleiotropic actions. The intent is to evaluate the likelihood of employing this substance for therapeutic purposes, for instance in diseases where a deficit could be part of the underlying pathophysiological mechanisms, also avoiding any adverse effect. Relaxin is already being considered as a promising drug, especially in acute heart failure. A careful study of the different RXFPs and their receptors and the comprehension of all biological activities of these hormones will probably provide new drugs with a potential wide range of therapeutic applications in the near future.
Cernaro V, Lacquaniti A, Lupica R, et al., Med Res Rev. 2014 Jan;34(1):77-105. doi: 10.1002/med.21277. Epub 2013 Feb 11.
Filippatos G, Teerlink JR, Farmakis D et al., Eur Heart J. 2013 Dec 6. [Epub ahead of print]
Alexiou K, Wilbring M, Matschke K, Dschietzig T., PLoS One. 2013 Sep 30;8(9):e75592. doi: 10.1371/journal.pone.0075592.
CONCLUSION: Relaxin improved endothelial dysfunction by promoting eNOS activity, suppressing endothelin-1 and arginase-II expression, and up-regulating SOD1 via GR, GR-c/EBP-β, and PI3K-Akt pathways. This corroborates the notion that it functions as an endogenous and potentially therapeutic vasoprotector.
The publication used the human relaxin 2 peptide from Phoenix Pharmaceuticals for investigation dose-dependent vasodilation-restoration by aortic ring vascular assays
Dschietzig T, Brecht A, Bartsch C, Baumann G, Stangl K, Alexiou K. Cardiovasc Res. 2012;95(1):97-107.
Chan LJ, Rosengren KJ, Layfield SL et al., J Biol Chem. 2012 Nov 30;287(49):41152-64. doi: 10.1074/jbc.M112.409284. Epub 2012 Sep 28.
Nascimento AR, Pimenta MT, Lucas TF, et al., Eur J Pharmacol. 2012 Sep 15;691(1-3):283-91. doi: 10.1016/j.ejphar.2012.07.021. Epub 2012 Jul 20.
Effect of Relaxin on Human Sperm Functions
Relaxin is a circulating hormone with functions in pregnancy, parturition, and other aspects of female reproduction. It is also secreted from the prostate gland into the seminal fluid; however, the role of relaxin in male reproduction is debated. Studies conducted in the past have suggested possible actions on human spermatozoa, but the data were contrasting. Here, we show that the relaxin receptor RXFP1 (Relaxin Family Peptide Receptor 1) is expressed in human spermatozoa, and it mainly localizes in the astrodome. In vitro studies on human sperm demonstrated that this hormone attenuates the natural decline in sperm motility and maintains higher mitochondrial activity and lower apoptosis level. Furthermore, relaxin induced an increase in sperm hyperactivation, intracellular calcium and cAMP, and acrosome reaction. These effects were abolished by the use of the specific anti-RXFP1 antibody. Relaxin concentrations were low in the blood (x¯ 6 SD, 0.16 6 0.03 nM) and very high in the seminal plasma (x¯ 6 SD, 10.3 6 4.0 nM), confirming its secretion mainly by the prostate. Taken together, these data demonstrate that relaxin influences positively many sperm functions linked to fertilizing ability, and it preserves sperm functionality, with possible practical value in assisted reproduction techniques.
This publication used Human relaxin-2 from Phoenix Pharmaceuticals for sperm function studies.
Ferlin A, Menegazzo M, Gianesello L, Selice R, Foresta C. Effect of relaxin on human sperm functions. J Androl. 2012;33(3):474-82.
Ferlin A, Menegazzo M, Gianesello L, et al., J Androl. 2012 May-Jun;33(3):474-82. doi: 10.2164/jandrol.110.012625. Epub 2011 Sep 8.
Hossain MA, Rosengren KJ, Samuel CS et al., J Biol Chem. 2011 Oct 28;286(43):37555-65. doi: 10.1074/jbc.M111.282194. Epub 2011 Aug 30.
McGowan BM, Minnion JS, Murphy KG et al., Diabetes Obes Metab. 2010 Dec;12(12):1090-6. doi: 10.1111/j.1463-1326.2010.01302.x.
This Publication used LGR7 and LGR8 antibodies from Phoenix Pharmaceuticals for IHC staining.
Filonzi M, Cardoso LC, Pimenta MT, et al. Reprod Biol Endocrinol. 2007;5:29.
In humans, the relaxin hormone family includes H1, H2 and H3 isoforms and insulin-like peptides 3 to 6. The ever-increasing interest in relaxin as potential new drug requires reliable methods to compare bioactivity of different relaxins. The existing bioassays include in vivo or ex vivo methods evaluating the organ-specific responses to relaxin and in vitro methods based on measurement of cAMP increase in relaxin receptor-bearing cells. We previously demonstrated that relaxin dose-dependently inhibits platelet aggregation. On this basis, we have developed a simple, reliable bioassay for relaxin used to compare purified porcine relaxin, assumed as reference standard, with two recombinant human H2 relaxins, H3 relaxin, insulin-like peptides 3 and 5. Pre-incubation of platelets with relaxins (3, 10, 30,100, 300 ng/ml; 10 min.) caused the inhibition of ADP-induced platelet aggregation. Within the 10-100 ng/ml range, porcine relaxin showed the highest effects and a nearly linear dose-response correlation. Lower peptide concentrations were ineffective, as were insulin-like peptides 3 and 5 at any concentration assayed. Platelet inhibition was mediated by specific RXFP1 relaxin receptor and cGMP, whose intracellular levels dose-dependently increased upon relaxin. For comparison, we stimulated THP-1 cells, a relaxin receptor-bearing cell line, with porcine relaxin, human H2 and H3 relaxins at the above concentrations (15 min.). We observed a dose-related increase of intracellular cAMP similar to the trend of platelet inhibition. Insulin like peptide 5 was ineffective. In conclusion, this study shows that inhibition of platelet aggregation may be used to assess bioactivity of relaxin preparations for experimental and clinical purposes.
This publication used human relaxin-2 peptide from Phoenix Pharmaceuticals for ADP-induced platelet aggregation study.
Bani D, Nistri S, Cinci L, et al. A novel, simple bioactivity assay for relaxin based on inhibition of platelet aggregation. Regul Pept. 2007;144(1-3):10-6.
Cosen-Binker LI, Binker MG, Cosen R et al., World J Gastroenterol. 2006 Mar 14;12(10):1558-68.
Relaxin peptides are important hormones for the regulation of reproductive tissue remodeling and the renal cardiovascular system during pregnancy. Recent studies demonstrated that two of the seven human relaxin family peptides, relaxin H2 (RLN2) and INSL3, signal exclusively through leucine-rich repeat-containing G protein-coupled receptors, LGR7 and LGR8. Although it was well characterized that an RXXXRXXI motif at the RLN2 B chain confers receptor activation activity, it is not clear what roles RLN2 A chain plays in receptor interaction. Analyses of relaxin family genes on syntenic regions of model tetrapods showed that the A chain of RLN2 orthologs exhibited a greater sequence divergence as compared with the receptor-binding domain-containing B chain, foreshadowing a potential role in receptorinteractions; hence, defining receptor selectivity in this fast evolving peptide hormone. To test our hypothesis that select residues in the human RLN2 A chain play key roles in receptor interaction, we studied mutant peptides with residue substitution(s) in the A chain. Here, we showed that alanine substitution at the A16 and A17 positions enhances LGR8-activation activity of RLN2, whereas mutation at the A22-23 region (RLN2A22-23) ablates LGR8, but not LGR7, activation activity. In addition, we demonstrated that the functional characteristics of the RLN2A22-23 mutant are mainly attributed to modifications at the PheA23 position. Taken together, our studies indicated that ThrA16, LysA17, and PheA23 constitute part of the receptor-binding interface of human RLN2, and that modification of these residues has led to the generation of novel human RLN2 analogs that would allow selective activation of human LGR7, but not LGR8, in vivo.
The publication used human relaxin 2 peptide tracer (Cat.#T-035-62) for receptor binding experiments
Park JI, Semyonov J, Yi W, Chang CL, Hsu SY. J Biol Chem. 2008;283(46):32099-109.
Effect of relaxin on myocardial ischemia injury induced by isoproterenol.
Zhang J, Qi YF, Geng B, et al., Peptides. 2005 Sep;26(9):1632-9.
Schematics













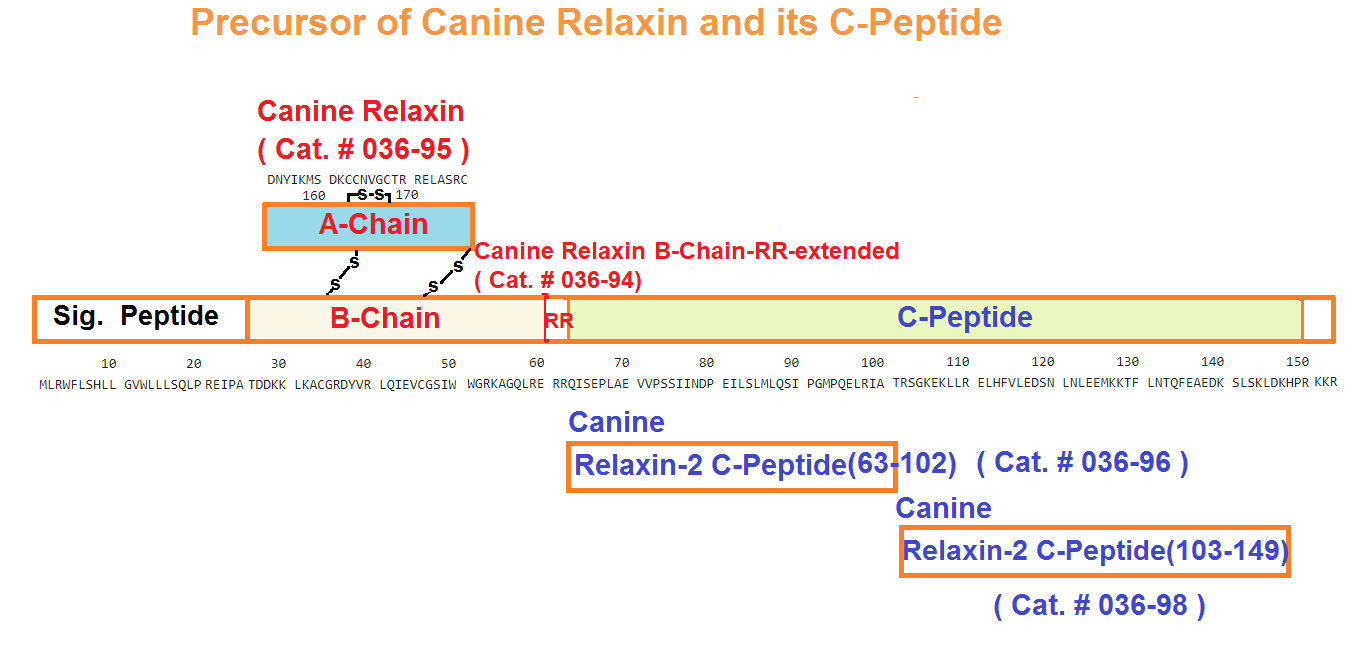
More Information

